![]()
![]()
![]()
![]()
![]()
![]()
![]()
Future Directions in Bioluminescence Research
ABSTRACT
Flow visualization in the ocean –
Implications of laboratory bioluminescence experiments
Jim Rohr1, Jon Schoonmaker1, Michael Latz2 and Mark Hyman3
1. SPAWARSYSCEN San Diego CA. 92152-5000
2. Scripps Institution of oceanography, UCSD, CA. 92093-0209
3. Coastal Systems Station, NSWC Panama City, Fl. 32407
 Bioluminescence is recognized to be one of the most cosmopolitan organism behaviors in the marine environment. It is considered so ubiquitous that its absence is now considered more remarkable than its presence. One of the primary sources of bioluminescence in all oceans are unicellular plankton, called dinoflagellates. In coastal waters where concentrations of dinoflagellates are greatest, opportunities to observe flow stimulated bioluminescence are typically best. It has been suggested that concentrations of less than 100 dinoflagellates/liter-are sufficient to highlight moving objects. Where extended time series monitoring of coastal bioluminescence have been conducted, it has been generally found that the numbers of bioluminescent dinoflagellates are normally equal to or greater than 100 cells l-1. During red tide blooms the abundance of luminescent dinoflagellates may reach 3 x 107 cell l-1.
Bioluminescence is recognized to be one of the most cosmopolitan organism behaviors in the marine environment. It is considered so ubiquitous that its absence is now considered more remarkable than its presence. One of the primary sources of bioluminescence in all oceans are unicellular plankton, called dinoflagellates. In coastal waters where concentrations of dinoflagellates are greatest, opportunities to observe flow stimulated bioluminescence are typically best. It has been suggested that concentrations of less than 100 dinoflagellates/liter-are sufficient to highlight moving objects. Where extended time series monitoring of coastal bioluminescence have been conducted, it has been generally found that the numbers of bioluminescent dinoflagellates are normally equal to or greater than 100 cells l-1. During red tide blooms the abundance of luminescent dinoflagellates may reach 3 x 107 cell l-1.
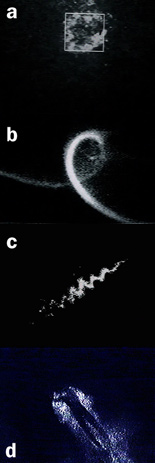
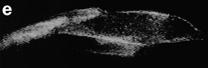
The video frames presented on this page were recorded with an intensified video camera and are illustrative of the bioluminescent displays visible to the naked eye, even when the flow stimuli are relatively weak. Proceeding from top-to-bottom are bioluminescent images of a 0.25-m diameter sphere (a) and a coiled rope (b), discernable several meters beneath the sea surface as a result of the bioluminescence stimulated by passing swell. The total bioluminescence flux from the 0.25-m diameter sphere was estimated to be about 2.9 x 1011 photons s-1 (Rohr et al., 1994). Also shown are the bioluminescent signatures of (c) a swimming fish, (d) diver, and (e) dolphin. Although flow-induced bioluminescent displays are common and often quite spectacular, before bioluminescence can be seriously considered as a possible strategy for nighttime flow visualization in the ocean, two issues must be addressed:
- Do luminescent organisms exhibit a characteristic and repeatable flash response to hydrodynamic agitation?
- Under what ambient light conditions can bioluminescence be detected?
Issue 1: The principal focus of our research was to study the luminescent response of freshly collected sea water samples of mixed plankton to repeatable and quantifiable levels of flow agitation. The flow chosen was fully developed pipe flow where the shear stress and turbulent length scales, believed to be important for hydrodynamic stimulation (e.g, Latz & Rohr, 1999), have been exhaustively studied. Pipe flow experiments were conducted in three different diameter pipes, which together provided shear stress levels (0.2 dynes cm-2 – 1.45x103 dynes cm-2) and turbulent length scales (3mm – 2.54x104mm) spanning nearly four orders of magnitude. Several recurring trends were found throughout all the pipe-flow data sets. These included a repeatable threshold level for bioluminescence stimulation that occurred in laminar flow at a shear stress of approximately 1-dyne cm-2 (see below for representative measurements).
 Maximum flash levels of individual cells also exhibited a conspicuous trend. After exhibiting a graded response with wall shear stress increasing from 1-10 dynes cm-2, maximum flash intensities remained essentially constant thereafter (tested up to wall shear stresses of 2000 dynes cm-2). This was true regardless of the laminar/turbulent nature of the flow, even as the turbulent length scales became an order of magnitude smaller than the size of the smallest luminescent dinoflagellates present. Together with a ship-wake numerical simulation, the pipe flow data sets provided a means for predicting the spatial extent of the bioluminescent "footprint" of the wake of an aircraft carrier. At 18 kts (9 m s-1) the stimulatory wake was estimated to be over 5 km long.
Maximum flash levels of individual cells also exhibited a conspicuous trend. After exhibiting a graded response with wall shear stress increasing from 1-10 dynes cm-2, maximum flash intensities remained essentially constant thereafter (tested up to wall shear stresses of 2000 dynes cm-2). This was true regardless of the laminar/turbulent nature of the flow, even as the turbulent length scales became an order of magnitude smaller than the size of the smallest luminescent dinoflagellates present. Together with a ship-wake numerical simulation, the pipe flow data sets provided a means for predicting the spatial extent of the bioluminescent "footprint" of the wake of an aircraft carrier. At 18 kts (9 m s-1) the stimulatory wake was estimated to be over 5 km long.
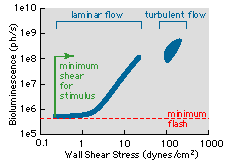 Threshold shear stress levels of the order of 1-dyne cm-2 are higher than what would normally be associated with flows within most of the ocean. This is in agreement with the general lack of "spontaneous" dinoflagellate bioluminescence observed. The stimulation of luminescent dinoflagellates at shear stress levels of about 1 dyn cm-2 in laminar flow is also consistent with observed occurrences of bioluminescence, e.g. breaking waves, swimming fish and its function as an antipredation strategy (Latz et al. 1994).
Threshold shear stress levels of the order of 1-dyne cm-2 are higher than what would normally be associated with flows within most of the ocean. This is in agreement with the general lack of "spontaneous" dinoflagellate bioluminescence observed. The stimulation of luminescent dinoflagellates at shear stress levels of about 1 dyn cm-2 in laminar flow is also consistent with observed occurrences of bioluminescence, e.g. breaking waves, swimming fish and its function as an antipredation strategy (Latz et al. 1994).
Issue 2: Because ambient night-light from the moon and stars at the ocean surface are often much brighter than bioluminescent light, it is necessary to develop effective strategies to extract bioluminescence signals from ambient noise. Over the last 20 years, land based multi- and hyper-spectral technology have progressed to the point where low contrast objects can be detected in cluttered backgrounds because of subtle differences between object and background spectra. Within the dynamic range of the sensor, bioluminescent dinoflagellates are excellent candidates for exploitation by multi-spectral technologies. Dinoflagellate spectra peak emission is around 470-480 nm (blue) with a half-maximum full width of about 50 nm (Losee and Lapota, 1981). In contrast, competing background sources of moonlight, starlight, airglow and zodiacal light are nearly all equally rich in the red as they are in the blue.
The images below serve as an example of the advantages offered by a simple multi-spectral approach. In the unprocessed image on the left (a), whales are barely discernable because light reflected at the sea surface obscures the upwelling light, which serves to silhouette whales within the water column. Generally, sea surface induced variance in the spectral bands of interest is much larger than any spectral variance introduced by the presence of the whale. However, because the spectra of reflected and upwelling light are different (in this simple analysis, green is only associated with reflected light), sea surface variance can be greatly reduced. The processed image on the right (b), which clearly shows the silhouettes of two whales, is achieved by simply subtracting a weighted green band from the blue band. This weight is determined by calculating the covariance between the blue and green bands (for further details see Barnes et al. 1999). For bioluminescence a similar approach can be taken. A correctly tuned multi-spectral sensor will have at least one target band in which the bioluminescence signature is detected and one to three guard bands, which correlate to the background clutter and other noise terms. 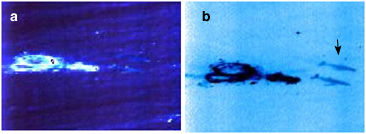
References
Barnes, C., G. Gilbert, J. Schoonmaker, and J. Rohr (1999) The possibility of passive whale tr4acking with the use of a hyperspectral sensor. Oceans ’99 MTS/IEEE, 13-16 Sept., Seattle Wash.
Latz, M.I., J.F. Case and R.L. Gran (1994) Excitation of bioluminescence by laminar shear associated with simple Couette flow. Limnol. Oceanogr. 39: 1424-1439.
Latz, M.I. and J. Rohr (1999) Luminescent response of the red tide dinoflagellate Lingulodinium polyedrum to laminar and turbulent flow, Limnol. and Oceanogr. 44(6), 1423-1435.
Losee, J. and D. Lapota (1981) Bioluminescence measurements in the Atlantic and Pacific. Pages 143-152 in K. Nealson [ed.], Bioluminescence: Current Perspectives.
Rohr, J., J. Losee and G. Anderson (1994) The response of bioluminescent organisms to fully developed pipe flow. Naval Command, Control and Ocean Surveillance Center, RDT&E Div. Tech. Rep. No.1360.
[ next abstract >> ]
[Download printable PDF version of this page]
 . | Return to SD Workshop Page. | Return to the Bioluminescence Web Page.
. | Return to SD Workshop Page. | Return to the Bioluminescence Web Page.